Als eine der wichtigsten Kulturpflanzen ist Mais weltweit bekannt und ruft in den meisten Köpfen wahrscheinlich ein ähnliches Bild hervor: eine verdickte Spindel, eng besetzt mit sattgelb gefärbten Samen (Maiskörner), welche von Blättern eng umhüllt und mit seidig schimmernden Fasern an der Spitze des Kolbens besetzt ist. Dies ist das Bild von hochmodernem, hochproduktivem Hybridmais, wie er heutzutage in einem großen Teil der Welt angebaut wird. Betrachtet man allerdings den Vorfahren von Mais, Teosinte, eine vor allem in Mittelamerika konzentrierte, wildwachsende Maisart werden auf den ersten Blick deutliche Unterschiede in der Pflanzen-architektur und der Zahl und Form der Ähren/Kolben klar (Abb. 1A). Als eine auskreuzende Art mit einem doppelten Chromosomensatz, führten die wiederkehrenden Effekte von Rekombination und das Einwirken evolutionärer Kräfte während der Domestizierung von Teosinte, die vor ca. 9000 Jahren begann, zu schrittweisen Veränderungen des Chromosomensatzes. Das erklärt die heutigen Unterschiede zwischen Teosinte und modernem Mais.
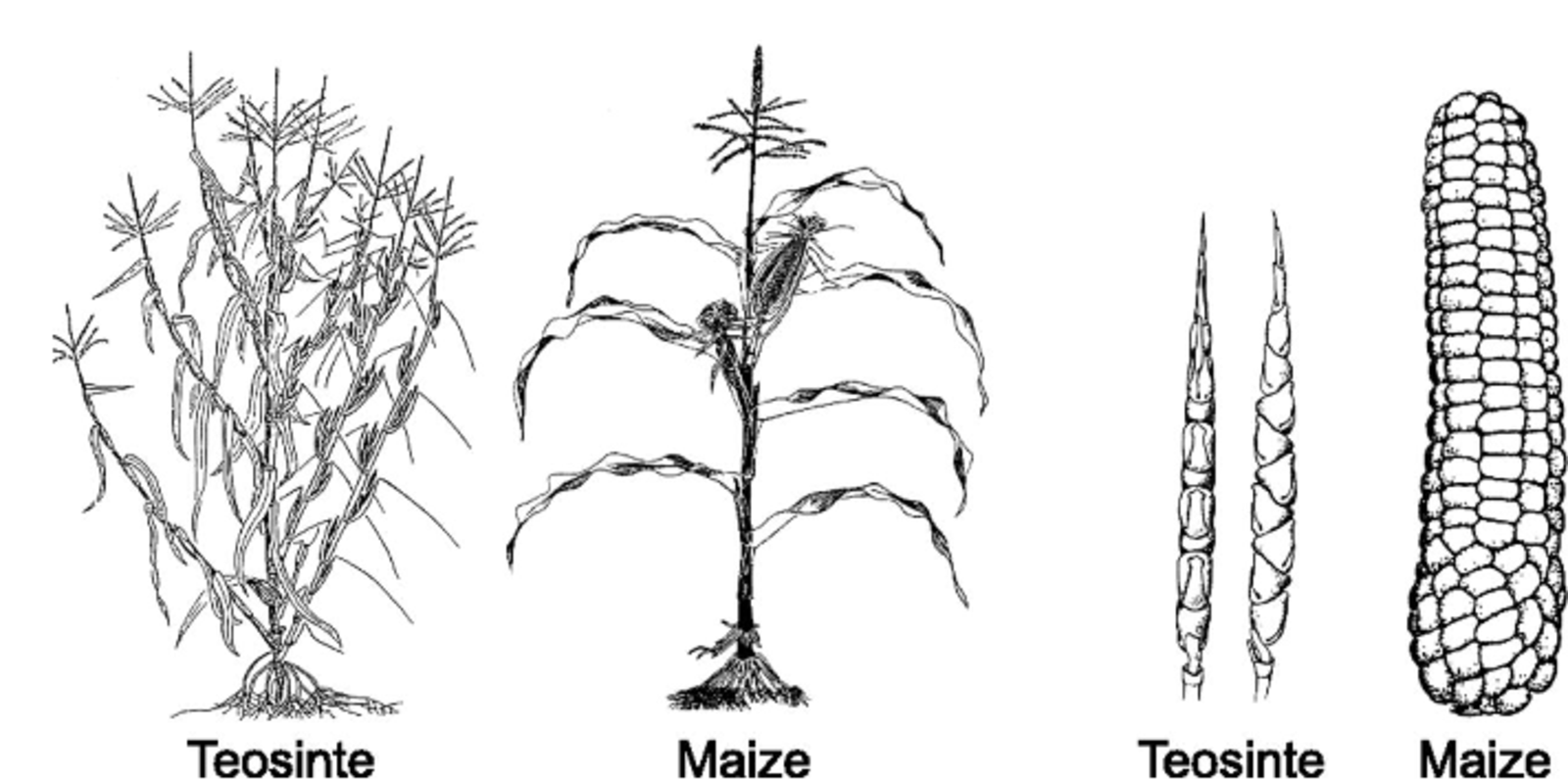
Der moderne Kulturmais wird heute weltweit angebaut, auch fast überall in Europa. Dennoch gibt es, wenn es um Mais geht, auch heute noch mehr Diversität, als man zunächst vermuten würde (Abb. 1B). Denn Mais hat eine komplizierte Geschichte, charakterisiert durch eine Abfolge demografischer Ereignisse, Populationsengpässe und -expansionen, Migration und Vermischung zwischen verschiedenen regionalen Populationen. Das hat zu einer Vielzahl verschiedener regionaler Maispopulationen mit diversen Genpools geführt. Allein in Europa gibt es hunderte lokaler Sorten, soge-nannte Landrassen, welche allein durch traditionelle landwirtschaftliche Methoden verbessert wurden. Bis in die 1960er Jahre wurden diese Sorten intensiv genutzt, bis sie durch den produktiveren, aber auch weniger diversen, modernen Mais verdrängt wurden. Heutzutage lagern diese europäischen Landrassen vorwiegend in Genbanken. Viele von diesen Landrassen sind besonders gut an ihre Standorte angepasst und besitzen zum Teil in der Pflanzenzüchtung gewünschte Eigenschaften in Form von seltenen Genvarianten, welche im modernen Mais mit seinem starken Züchtungsfokus auf Ertrag verloren gegangen sind. Der Ursprung und die Geschichte dieser Genvarianten sind heutzutage noch unklar. Wann sind sie erstmals aufgetaucht? Waren sie bereits in Teosinte vorhanden? Wie gelangten sie nach Europa? Oder entstanden sie erst, als Mais um 1492 erstmals nach Europa gebracht wurde?

Um dies herausfinden zu können, müssen wir den Maisgen-Varianten in die Vergangenheit folgen. Der Grundgedanke dieser Idee basiert auf der grundlegenden Vorstellung, dass die Abstammungsbeziehungen aller Maisindividuen theoretisch durch einen einzigen Stammbaum beschrieben werden können. Diploide, sich sexuell fortpflanzende Organismen, zu denen Mais gehört, erhalten einen Chromosomensatz von ihrer ”Mutter” und einen von ihrem ”Vater”, welche ihrerseits jeweils einen Chromosomensatz von ihren beiden Eltern erhalten haben und so weiter. Diese Art der Vererbung von genetischem Material ermöglicht es, die Abstammungsbeziehungen zwischen Individuen als einen Stammbaum zu visualisieren. Diese Darstellung funktioniert sowohl zwischen Individuen als auch zwischen DNA-Sequenzen. Ein Baum beschreibt die Abstammung einer DNA-Sequenz jedoch nur dann genau, wenn diese als Einheit (Haplotyp) über Generationen weitergegeben wird. Die Neuanordnung von genetischem Material (Rekombination) während der Keimzellenbildung (Meiose) teilt DNA-Segmente und bricht diese Einheiten auf. Während man das Genom durchquert, haben verschiedene Teile des Genoms zunehmend unterschiedliche Vorfahren und müssen anhand verschiedener Bäume beschrieben werden (Abb. 2a). Daraus resultiert ein Stammbaumnetzwerk, dass alle Ahnenbeziehungen innerhalb einer Sammlung von Chromosomensätzen von einer Vielzahl von Individuen darstellt (Abb. 2b).
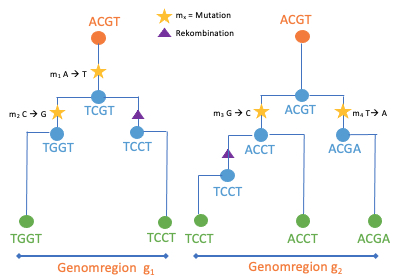
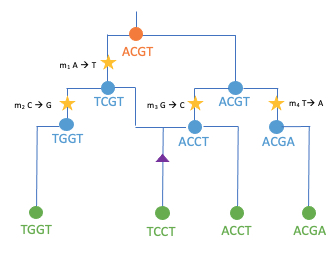
Abb. 2 Schematische Übersichten der gemeinsamen Abstammung.
a.) Stammbaum, der beschreibt, wie eine Ansammlung von Chromosomensätzen an einer chromosomalen Stelle miteinander verwandt ist.
b.) Stammbaumnetzwerk, die Zusammenfassung einer Sequenz korrelierter Bäume, wenn man den Genbäumen entlang eines Genoms folgt [inspiriert von [3, 4, 5]]
Ziel meiner Arbeit ist es, mit Hilfe neuer Algorithmen aus einer Sammlung von Chromosomensätzen einer Vielzahl von Maisindividuen, den DNA-Segmenten naher Verwandter und der Wildform von Mais (Teosinte) alle dazwischenliegenden DNA-Segmente abzuleiten und damit das gesamte Stammbaumnetzwerk für Mais zu rekonstruieren. Sobald mir dies gelungen ist, werde ich Genvarianten von Interesse innerhalb dieses Stammbaum-Netzwerkes in die Vergangenheit folgen, ihr erstes Auftauchen datieren und ihre geografischen Standorte und historischen Migrationen bestimmen und diese mit der bekannten Geschichte von Mais abgleichen. Dies würde uns einen bis jetzt unerreicht umfassenden Einblick in die Geschichte und Evolution von Mais ermöglichen. All dies kann uns helfen zu verstehen, wie Pflanzen sich an ihre Umwelt anpassen. Es würde uns zudem zeigen, welche Genvarianten für diese Anpassung in Mais besonders wichtig waren und uns erlauben diese Genvarianten auch wieder in modernem Mais zu integrieren.
Planter's Punch
Unter der Rubrik Planter’s Punch wird jeden Monat ein bestimmter Aspekt des CEPLAS Forschungsprogramms vorgestellt. Alle Beiträge werden von Mitgliedern der Graduiertenschule und des Postdoc Programms erstellt.
Über die Autorin

Kerstin Schulz arbeitet an der Domestizierungs- und Kolonisationsgeschichte von Mais unter Verwendung von Ahnenrekombinationsgraphen. Dazu versucht sie Alter, Herkunft und geographische Verteilung aller Maisallele im europäischen Mais abzuleiten. Ihr Ziel ist es, die Evolutionsgeschichte verschiedener Teosinte- und Maispopulationen entlang der Expansionsroute von Mais besser zu verstehen und die populationsgenetischen Signale zu erkennen, welche die genetische Diversität von Mais geprägt haben.
Bildernachweise
[1] Chin Jian Yang et al. “The genetic architecture of teosinte catalyzed and constrained maize domestication”. In: Proceedings of the National Academy of Sciences 116.12 (2019), pp. 5643–5652.
[2] CIMMYT Maize Germplasm Bank. Maize: From Mexico to the world.